Zameranie na systém renín-angiotenzín-aldosterón na prevenciu hypertenzie a ochorenia obličiek vývojového pôvodu
Mar 24, 2022
Kontakt: Audrey Hu Whatsapp/hp: 0086 13880143964 E-mail:audrey.hu@wecistanche.com
Chien-Ning Hsu1,2a You-Lin Tain3,4,*
Abstrakt:
Systém renín-angiotenzín-aldosterón (RAAS) sa podieľa na hypertenzii a ochorení obličiek. Vyvíjajúca sa oblička môže byť naprogramovaná rôznymi rannými poruchami takzvaným obličkovým programovaním, čo vedie k hypertenzii a ochoreniu obličiek v dospelosti. Táto teória je známa ako vývojový pôvod zdravia a choroby (DOHaD). Naopak, skoré intervencie založené na RAAS by mohli zvrátiť programové procesy, aby sa predišlo výskytu choroby takzvaným preprogramovaním. V tomto prehľade sumarizujeme hlavne (1) súčasné poznatky o RAAS, ktoré sa podieľajú na programovaní obličiek; (2) súčasné dôkazy podporujúce spojenia medzi aberantným RAAS a inými mechanizmami za programovaním obličiek, ako je oxidačný stres, nedostatok oxidu dusnatého, epigenetická regulácia a dysbióza črevnej mikroflóry; a (3) prehľad o tom, ako môžu zásahy preprogramovania založené na RAAS zabrániť hypertenzii a ochoreniu obličiek vývojového pôvodu. Na urýchlenie prechodu intervencií založených na RAAS na prevenciu hypertenzie a ochorenia obličiek je potrebné rozšírené pochopenie RAAS, ktoré sa podieľa na programovaní obličiek, ako aj väčšie zameranie na ďalší klinický preklad.
Kľúčové slová:chronické ochorenie obličiek; hypertenzia; systém renín-angiotenzín-aldosterón; oxid dusnatý; vývojový pôvod zdravia a choroby (DOHaD); oxidačný stres; angiotenzín-konvertujúci enzým; nefrón

Cistanche deserticola zabraňuje ochoreniu obličiek
1. Úvod
Hypertenzia a chronické ochorenie obličiek (CKD) sú vysoko rozšírené ochorenia na celom svete. WHO uvádza, že jeden zo štyroch mužov a jedna z piatich žien má hypertenziu [1]. CKD postihuje až desať percent svetovej populácie [2]. Hypertenzia a CKD sú úzko prepojené [3], takže CKD je jednou z najčastejších príčin sekundárnej hypertenzie a hypertenzia je dôležitým faktorom súvisiacim s progresiou CKD. Najznámejším príkladom je stenóza renálnej artérie, ktorá je charakterizovaná hypertenziou a progresívnou stratou renálnej funkcie [4]. Bol uznaný ako prototyp angiotenzín-dependentnej hypertenzie, čo prispelo k objavu systému renín-angiotenzín-aldosterón (RAAS) [5].
Rastúci počet dôkazov naznačuje, že hypertenzia aj ochorenie obličiek môžu mať svoj pôvod v ranom veku [6–8]. Expozícia suboptimálnemu vnútromaternicovému prostrediu počas vývoja obličiek vedie k celoživotným negatívnym vplyvom na obličkovú štruktúru a funkciu a na obličkové kompenzačné mechanizmy takzvaným renálnym programovaním [9,10]. Vyvíjajúca sa oblička môže byť naprogramovaná rôznymi rannými ochoreniami, čo vedie k hypertenzii a ochoreniu obličiek v dospelosti. Koncept, že nepriaznivé podmienky počas organogenézy zvyšujú náchylnosť na rozvoj chorôb dospelých, sa nazýva hypotéza fetálneho pôvodu [11], novšie pomenovaná ako „Developmental Origins of Health and Disease“ (DOHaD) [12]. Na druhej strane tento koncept vedie k teoretickému posunu terapeutického prístupu z dospelého života do skoršieho štádia, konkrétne preprogramovania, s cieľom potenciálne zvrátiť chorobné procesy skôr, ako sa klinické ochorenie prejaví [13,14].
Krvný tlak (BP) je prísne kontrolovaný veľmi zložitými sieťami, vrátane RAAS, endotelovej funkcie, sympatického nervového systému, natriuretických peptidov, zápalu a imunitného systému [15–17]. RAAS má kontraregulačnú úlohu v patogenéze a rozvoji hypertenzie [17]. Bolo riešených niekoľko potenciálnych molekulárnych mechanizmov zapojených do vývojového programovania hypertenzie a ochorenia obličiek, vrátane aberantného RAAS, oxidačného stresu, nedostatku oxidu dusnatého (NO), dysbiózy črevnej mikroflóry, dysregulovaných signálov snímania živín, epigenetickej regulácie a zníženého počtu nefrónov [ 6–9,13,14,18–20]. Spomedzi nich hrá RAAS nielen zásadnú úlohu pri regulácii BP, ale úzko spolupracuje aj s inými mechanizmami. RAAS je hlavná hormonálna kaskáda zložená z rôznych angiotenzínových peptidov s rôznymi biologickými funkciami sprostredkovanými odlišnými receptormi [21]. V RAAS existujú dve hlavné cesty: klasické a neklasické cesty. Klasický RAAS sa skladá hlavne z enzýmu konvertujúceho angiotenzín (ACE), angiotenzínu (ANG) II a receptora angiotenzínu II typu 1 (AT1R). Za patofyziologických podmienok môže byť klasický RAAS aktivovaný na spustenie vazokonstrikcie a zápalu, čím sa podporuje hypertenzia a poškodenie obličiek [22]. Naopak, neklasický RAAS zložený z osi receptora ACE2-ANG-(1-7)-MAS vyvažuje škodlivé účinky signalizácie ANG II.
Je potrebné poznamenať, že obe osi RAAS sú spojené s programovaním plodu [23,24]. Hoci blokáda klasického RAAS poskytuje základ pre súčasné antihypertenzívne a renoprotektívne terapie [25], existujú obmedzené údaje o tom, či včasné zacielenie na RAAS môže zabrániť hypertenzii a ochoreniu obličiek vývojového pôvodu.
V prehľade preto uvádzame súčasnú aktualizáciu RAAS, vysvetľujúc jeho úlohu pri hypertenzii a chorobách obličiek vývojového pôvodu a zdôrazňujúcich jeho väzby na iné mechanizmy. Zdôrazňujeme tiež potenciálne preprogramovacie zásahy, ktoré sa zameriavajú na RAAS na prevenciu vývojového programovania hypertenzie a ochorenia obličiek. Získali sme súvisiacu literatúru zo všetkých článkov indexovaných v PubMed/MEDLINE. Použili sme tieto kľúčové slová a ich kombinácie: "renín", "angiotenzín", "chronické ochorenie obličiek", "vývojové programovanie", "DOHaD", "potomstvo", "matka", "nefrogenéza", "nefrón", "prorenín" "receptor", "aldosterón", "mineralokortikoidný receptor", "tehotenstvo", "potomstvo", "preprogramovanie", "angiotenzinogén", "angiotenzín-konvertujúci enzým" a "hypertenzia". Ďalšie štúdie sa potom vybrali a vyhodnotili na základe vhodných referencií vo vhodných dokumentoch. Posledné vyhľadávanie sa uskutočnilo 30. januára 2021.

Výhody extraktu z cistanche tubolosa:liečenie ochorení obličiek
2. RAAS a programovaná oblička
2.1. Intrarenálny RAAS
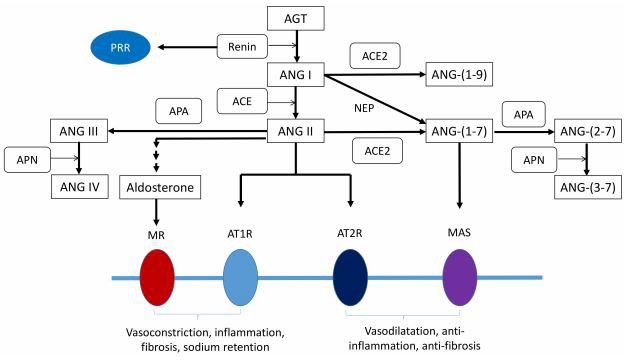
Obličky sú hlavným cieľom pre rôzne zložky RAAS, ktoré zahŕňajú prorenín/renín, ANG II, ANG III (ANG-(2–8)), ANG-(1–7), ANG IV (ANG-(3– 8)), ANG-(1–9) a aldosterón [26]. Renin spustí kaskádu udalostí v RAAS. Obličky sú jediným známym orgánom, kde dochádza ku konverzii prorenínu na renín [27]. Substrát RAS, angiotenzinogén (AGT) sa uvoľňuje z pečene a štiepi sa renínom, čím vzniká ANG I. ACE sa všeobecne vyskytuje v mnohých typoch buniek a tkanivách/orgánoch. ACE je primárne známy svojou schopnosťou štiepiť ANG I na ANG II, pričom štiepi nielen ANG I, ale aj mnohé ďalšie substráty vrátane bradykinínu [28]. ANG II stimuluje AT1R na zvýšenie reabsorpcie sodíka a zvýšenie TK [29]. Naopak, receptor ANG II typu 2 (AT2R) je ďalším typom receptora ANG II, ktorý sprostredkúva vazodilatáciu. V kôre nadobličiek pôsobí ANG II tak, že spôsobuje uvoľňovanie aldosterónu. Aldosterón podporuje retenciu sodíka stimuláciou transportéra sodíka v distálnych tubuloch obličiek, a preto zvyšuje TK. Je potrebné poznamenať, že renálny RAAS je charakterizovaný najvyššími tkanivovými koncentráciami ANG II [30]. V obličkách môže byť ANG II tiež metabolizovaný na ANG III (ANG-(2–8)) aminopeptidázou A (APA). Na druhej strane sa ANG III spracuje na ANG IV (ANG-(3–8)) aminopeptidázou N (APN) [30].
Na druhej strane ACE2, homológ ACE, konvertuje ANG II na ANG- (1–7) alebo konvertuje ANG I na ANG-(1–9) [31]. ANG-(1–7) indukuje natriuretické a diuretické účinky v prospech vazodilatácie prostredníctvom MAS receptora [24]. ANG I môže byť tiež premenený na ANG-(1–7) endopeptidázou neprilyzín (NEP) [30]. Na druhej strane, ANG-(1–7) môže byť spracovaný na (ANG-(2–7)) pomocou APA a ďalej metabolizovaný pomocou APN za vzniku ANG-(3–7) [30]. Hoci sa väčšina štúdií RAS zamerala hlavne na ANG II, ďalšie peptidové fragmenty Ang-(1-7), ANG III (ANG-(2–8)), ANG IV (ANG-(3–8)), ANG-(2–7) a ANG-(3–7) boli identifikované ako potenciálne bioaktívne [30]. Keďže rôzne peptidy v RAAS môžu fungovať v zhode alebo v opozícii a že farmakologické zmeny RAAS vedú k súčasným zmenám rôznych ANG peptidov a kompenzačným zmenám v množstve/aktivite zúčastnených enzýmov RAAS, je potrebná rozsiahlejšia výskumná práca. pochopiť zložitosť siete peptidov RAAS a ako tento sieťový systém ovplyvňuje programovanie obličiek. Spracovanie rôznych ANG peptidov v RAAS v obličkách je znázornené na obrázku 1
2.2. Programovaná oblička: Príčina hypertenzie a ochorenia obličiek u dospelých?
Ľudské obličky sa skladajú z nefrónov v rozsahu od 250,{1}} do 1,1 milióna na obličku [32]. Nefrón je funkčnou jednotkou obličky, existuje však široká variabilita s 10-násobným individuálnym rozdielom [32]. Tvorba nefrónov, konkrétne nefrogenéza, začína v 9. a pokračuje až do 36. týždňa tehotenstva u ľudí [33]. K iniciácii vývoja obličiek dochádza, keď vyrastanie ureterického pupenu z nefrického kanálika napadne skupinu mezenchymálnych buniek obsiahnutých v kaudálnom konci nefrického povrazca. Vypracovanie ureterického pupene je známe ako vetviaca morfogenéza [34], ktorá vedie k tvorbe nefrónov a hromadeniu moču.
systém. Kľúčovým regulátorom rastu a vetvenia primárneho močovodu je neurotrofický faktor odvodený od gliálnych buniek (GDNF) [35]. Progenitory nefrónov sa epitelizujú a vytvárajú obličkové vezikuly, ktoré sa predlžujú na telo v tvare písmena S a potom sa úplne vyvinú do nefrónu. Medzi 18. a 32. týždňom dochádza k exponenciálnemu nárastu nefrónov. Počas tretieho trimestra je vývoj nefrónu ukončený medzi 32. a 36. týždňom gestácie [32]. V súlade s tým je normálne nefrogenéza dokončená v termíne. Predčasne narodené deti majú teda pri narodení pravdepodobne zníženú výbavu nefrónov. Počet nefrónov u predčasne narodených detí však závisí nielen od gestačného veku, ale aj od vnútromaternicového prostredia a perinatálnej starostlivosti. Zhoršená morfogenéza vetvenia môže spôsobiť nízku zásobu nefrónov a širokú škálu renálnych porúch, konkrétne vrodené anomálie obličiek a močových ciest (CAKUT).
Dôležitú podporu pre programovanie obličiek priniesla holandská kohortová štúdia hladomorových narodení, ktorá odhalila, že podvýživa počas tehotenstva má dlhodobé následky na zdravie dospelých, vrátane hypertenzie a ochorenia obličiek [36,37]. Niekoľko epidemiologických štúdií spájalo predčasnosť a nízku pôrodnú hmotnosť ako rizikové faktory pre ochorenie obličiek a hypertenziu v neskoršom veku [38–40]. Nízka pôrodná hmotnosť môže byť výsledkom obmedzenia vnútromaternicového rastu (IUGR) alebo predčasného pôrodu spojeného s nízkym počtom nefrónov [32,33,41]. Znížený počet nefrónov vedie ku kompenzačnej glomerulárnej hyperfiltrácii a glomerulárnej hypertenzii. Toto začína začarovaný kruh s ďalšou stratou nefrónov, ktorá vedie k zvýšeniu TK, poklesu funkcie obličiek a môže skončiť CKD.
Napriek tomu nie je možné určiť počet nefrónov u živých ľudí. Hoci použitie nanočastíc na báze feritínu ako cieleného kontrastného činidla pre magnetickú rezonanciu (MRI) na meranie počtu nefrónov v ľudských obličkách dosiahlo určitý pokrok [42], väčšiu pozornosť si zasluhuje validácia metódy na neinvazívne hodnotenie in vivo nefrónov. .
2.3. Vplyv RAAS na programovanie obličiek
Vo vyvíjajúcej sa obličke sú zložky RAAS vysoko exprimované a zohrávajú kľúčovú úlohu pri sprostredkovaní správnej morfológie a fyziologickej funkcie obličiek [43,44]. U potkanov môžu byť všetky zložky RAAS detegované v embryonálnych obličkách od 12. do 17. dňa gravidity, pričom u plodov a novorodencov potkanov sú vyššie hodnoty ako u dospelých potkanov [44]. U ľudí sa tehotným ženám vyhýbali lieky interferujúce s RAAS (napr. ACE inhibítory [ACEI] alebo blokátory angiotenzínových receptorov [ARB]) z dôvodu ACEI/ARB fetopatie a nesprávneho vývoja obličiek [45]. Predčasnosť bola spojená so zvýšením plazmatických hladín renínu a ANG II, ako aj aktivity ACE [46]. U zvierat, ktorým chýbajú gény RAAS, sa vyvinú výrazne abnormálne obličky [47,48]. Na druhej strane zvieratá transgénne pre gény RAS vykazujú hypertenziu [49]. Blokáda RAAS pomocou ARB losartanu počas 1. až 12. dňa postnatálneho života u potkana (počas štádia nefrogenézy) spôsobuje znížený počet nefrónov a hypertenziu v dospelosti [50].
Niektoré rizikové faktory pre rozvoj hypertenzie a ochorenia obličiek boli hodnotené v štúdiách na ľuďoch. Napriek tomu tieto pozorovacie štúdie nemôžu samy o sebe priamo stanoviť príčinnú súvislosť medzi rannými urážkami a ochorením dospelých. Okrem toho tieto štúdie na ľuďoch neosvetľujú molekulárne mechanizmy, ktorými sa vytvára hypertenzia a ochorenie obličiek, a poskytujú stratégiu preprogramovania. V dôsledku etických úvah týkajúcich sa toho, čo je možné alebo nie v štúdiách na ľuďoch, majú zvieracie modely veľký význam. Vzhľadom na to, že štúdie na ľuďoch majú veľa obmedzení, boli vytvorené zvieracie modely, aby sa preskúmali typy urážok, ktoré riadia programovanie obličiek, potenciálne mechanizmy programovania obličiek, zraniteľné obdobia počas vývoja obličiek a potenciálne stratégie preprogramovania.

extrakt z cistanche tubolosa: liečba ochorení obličiek
3. Zvieracie modely programovania obličiek: Vplyv RAAS
3.1. Renálne programovanie súvisiace s RAAS na zvieracích modeloch
V súčasnosti sa vytvára čoraz väčší počet zvieracích modelov na štúdium hypertenzie a ochorenia obličiek v rámci vývojového programovania. Ako je uvedené inde [6–10,13,14,18–20], niekoľko environmentálnych vplyvov v ranom veku, ktoré môžu naprogramovať obličky, čo vedie k hypertenzii a ochoreniu obličiek v neskoršom veku, ako je podvýživa matky, choroba matiek, fajčenie matiek a vystavenie liekom alebo toxínom z prostredia. Tabuľka 1 sumarizuje štúdie na zvieratách, ktoré preukazujú súvislosť medzi aberantným RAAS, rannými ochoreniami a následnou hypertenziou a ochorením obličiek u dospelých potomkov [51–88]. Tento prehľad sa obmedzuje iba na environmentálne urážky, ktoré sa vyskytujú počas trvania nefrogenézy, so zameraním na programovanie obličiek súvisiace s RAAS.
V tomto prehľade sa živočíšne druhy pohybujú od potkanov [51–60,62,63,65–69,71,73–77,79,80,82–87], myší [61,72,78,81], králikov [ 70] a ovce [64,88]. Potkany a myši boli dominantnými živočíšnymi druhmi používanými vo výskume na štúdium hypertenzie a ochorení obličiek vývojového pôvodu. Na rozdiel od ľudskej nefrogenézy, ktorá je dokončená in utero, pokračuje vývoj obličiek u hlodavcov až 2 týždne po narodení [89]. V súlade s tým môžu faktory prostredia nielen počas gravidity, ale aj v ranom období laktácie narušiť vývoj obličiek u hlodavcov, čo vedie k programovaniu obličiek a ochoreniu obličiek u dospelých. Tabuľka 1 ukazuje výsledky hodnotené u potkanov vo veku od 4 do 90 týždňov. Keďže jeden ľudský rok sa rovná dvom potkaním týždňom v dospelosti [90], väčšina hodnotených výsledkov sa rovná ľudskému veku od detstva po strednú dospelosť. Napriek tomu v podstate neexistujú žiadne informácie o veľkých zvieratách na štúdium vplyvu RAAS na hypertenziu a ochorenie obličiek vývojového pôvodu.
Tabuľka 1 uvádza, že podvýživa matiek je najčastejším faktorom súvisiacim s ochorením obličiek a hypertenziou vývojového pôvodu. Naprogramovanie obličiek môže spôsobiť celý rad nutričných porúch, vrátane vysokej spotreby sacharózy [51], diéty s vysokým obsahom fruktózy [52,53], obmedzenia bielkovín [58–63], obmedzenia kalórií [64], diéty s vysokým obsahom tukov [65,66 ], diéta s vysokým obsahom soli [67] a nízky príjem soli [68]. Po druhé, choroba matiek tiež zasahuje do programovania obličiek. Medzi tieto zdravotné stavy počas tehotenstva patrí hypertenzia [69,70], CKD [71], diabetes [72,73], chronodisrupcia [74], preeklampsia [75], infekcia [76], insuficiencia placenty [77] a hypoxia. [78]. Ďalším faktorom narúšajúcim programovanie obličiek je vystavenie chemikáliám alebo toxínom v životnom prostredí, ako je fajčenie [79,80], kofeín [81] a 2,3,7,8- tetrachlórdibenzo-p-dioxín (TCDD) [82]. . Okrem toho, programovanie obličiek môže byť spustené liekmi, ako je pyrolidínditiokarbamát [83] alebo glukokortikoid [84–88].
Najčastejším nežiaducim renálnym výsledkom sledovaného programovania obličiek je hypertenzia [51–65,67–88]. Albuminúria bola preukázaná u potomkov narodených matkám s obmedzením bielkovín [58], diabetom [72] alebo hypoxiou [78]. Rýchlosť glomerulárnej filtrácie (GFR), index renálnej funkcie, bola hlásená ako znížená [66,76], nezmenená [54,55,59,60] alebo dokonca zvýšená [58] v rôznych modeloch programovania obličiek. Okrem toho sú hlavnými morfologickými deficitmi znížený počet nefrónov [57,76,78,79], renálna hypertrofia [71], glomerulárna hypertrofia [78] a tubulointersticiálne poškodenie [66,78]. Tieto pozorovania naznačujú, že programovanie obličiek sa nespolieha na jeden konkrétny faktor a vykazuje širokú škálu fenotypov.
 in animal models. Studies tabulated according to animal models, species, and age at evaluation.")
3.2. Renín, (Pro)renín a ich receptor v obličkovom programovaní
Nežiaduce renálne výsledky súvisia so zvýšenou expresiou renínu [53,74,75,78,81,83,85,86] a/alebo PRR [74,85] vo väčšine, ale nie vo všetkých zvieracích modeloch (pozri tabuľku 1 ). Renín je syntetizovaný hlavne juxtaglomerulárnymi bunkami, ktoré sa nachádzajú v aferentných arteriolách obličiek ako preprorenín [91]. Signálny peptid sa počas prenosu odštiepi a vytvorí prorenín. Štiepením 43-aminokyselinového N-koncového fragmentu sa potom prorenín premení na aktívny renín. Oblička vylučuje do obehu renín aj prorenín. Okrem štiepenia AGT za vzniku ANG I sa renín viaže na PRR. Tento receptor tiež viaže prorenín. Proteín PRR je kódovaný pre Atp6ap2 (ATPáza 6 doplnkový proteín 2). Proteín PRR existuje v troch formách: (1) 35-39 kDa forma s plnou dĺžkou pozostávajúca z 3 domén, (2) 28 kDa rozpustná forma a (3) skrátená forma [91].
Keďže kaskáda RAAS začína renínom, vyvoláva to otázku, či môže blokovať RAAS v bode jeho aktivácie (tj renín), aby sa zabránilo programovaniu obličiek? Prvý selektívny inhibítor renínu, aliskiren, nie je horší ako ACEI a ARB pri znižovaní TK a bol hodnotený ako účinný antihypertenzívny liek [92]. Aliskiren inhibuje renín väzbou na jeho katalytické miesto, čím inhibuje aktivitu renínu a prorenínu, čím blokuje RAS. Hladiny renínu a prorenínu však zostávajú vysoké, čo by mohlo pravdepodobne indukovať signalizáciu PRR spôsobom nezávislým od ANG II.
V súčasnosti bola PRR identifikovaná pre svoje multifunkčné aspekty, vrátane (1) PRR zvyšuje RAAS katalyzovaním produkcie ANG I, (2) PRR indukuje signálnu dráhu mitogénom aktivovaných proteínkináz (MAPK), (3) PRR je potrebná ako podjednotku V-ATPázy, ktorá transportuje protóny cez plazmatickú membránu, a (4) PRR interaguje s kanonickými Wnt/-katenínovými a nekanonickými dráhami Wnt/planárnej bunkovej polarity (PCP), ktoré sú nevyhnutné pre embryonálny vývoj [91 93,94].
Mnohé správy ukázali, že signálna dráha PRR môže indukovať hypertenziu závislú od ANG II [91]. Zvieratá s nadmernou expresiou prorenínu vykazovali závažnú hypertenziu [95]. Na rozdiel od iných komponentov RAS sú PRR knockout myši letálne alebo dokonca tkanivovo špecifické a majú krátku očakávanú dĺžku života [96], čo naznačuje kľúčovú funkciu PRR, ktorá je nezávislá od (pro)renínu. V súčasnosti však existuje len málo dôkazov o úlohe dráhy prenosu signálu PRR nezávislej od ANG II na programovanú hypertenziu.
Naša predchádzajúca správa ukázala, že prenatálne podávanie dexametazónu (DEX) zvýšilo expresiu mRNA renínu (násobná zmena=2.41) a PRR (násobná zmena=2.37) počas štádia nefrogenézy [85]. Zvýšenie expresie renínu bolo pretrvávajúce až do veku 4 mesiacov a bolo spojené so zvýšeným TK, čo naznačuje vplyv PRR na programovanú hypertenziu vyvolanú DEX. Ďalej sme pozorovali, že vysoká hladina fruktózy u matky zvýšila expresiu renálneho renínu od 1 dňa (násobná zmena=3,05) do 3 mesiacov (násobná zmena=3,38) veku [97]. Tieto zistenia sú v súlade s predchádzajúcimi štúdiami, ktoré ukazujú zvýšenie aktivity plazmatického renínu u potomkov v rôznych programovacích modeloch [62,63,68]. V súčasnosti existuje málo spoľahlivých informácií, pokiaľ ide o proteín PRR a jeho následnú signalizáciu na zvieracích modeloch programovania obličiek. Či by znížená expresia PRR mohla vysvetliť absenciu účinkov závislých od PRR počas inhibície RAS, je potrebné ďalej objasniť [94]. Ďalej sme pozorovali, že následné signálne dráhy signálnych dráh PRR, MAPK a Wnt boli identifikované ako významné dráhy Kjótskej encyklopédie génov a genómov (KEGG) v obličkách potomstva pomocou sekvenovania RNA novej generácie v modeli inhibície NO [ 98]. Všetky tieto zistenia naznačujú, že dráha PRR môže byť terapeutickým cieľom pre programovanú hypertenziu.
Regulačné cesty súvisiace s PRR v rôznych programovacích modeloch sú znázornené na obrázku 2.
renin–PRR pathway is linked to programmed hypertension and kidney disease via ANG II-dependent and –independent effects.")
3.3. Klasická os RAAS v programovaní obličiek
Existujú konfliktné výsledky týkajúce sa up- a downregulácie klasických komponentov RAAS (tabuľka 1), z veľkej časti kvôli širokému vekovému rozpätiu, v ktorom sa hodnotili potomkovia. Vo väčšine štúdií sa u dospelých potomkov vyvinula hypertenzia a ochorenie obličiek, ktoré sa zhodovalo so zvýšenou expresiou ACE [64–66,72,74,76,82,86] a AT1R [51,54,55,59 ,62,63,67,69,72–74,78–81,84] a aktivita ACE [73,77,88].
Veľmi málo štúdií skúmalo RAAS v spojení s programovaním obličiek v rôznych vývojových štádiách. V modeli potkanov s nízkym obsahom bielkovín u matky [56] bola renálna expresia AT1R pri narodení potlačená, zatiaľ čo jej expresia bola zvýšená vo veku 4 týždňov. V inom modeli renálneho programovania vyvolanom placentárnou insuficienciou u potkanov Sprague Dawley sa u dospelých potomkov vyvinula hypertenzia v spojení so zvýšeným renínom a AGT mRNA, ako aj so zvýšenou aktivitou ACE vo veku 16 týždňov [77]. Naopak, expresia renínu a AGT mRNA bola pri narodení znížená [77]. Celkovo tieto zistenia v modeloch obličkového programovania naznačujú prechodnú bifázickú reakciu s downreguláciou klasických komponentov RAAS v novorodeneckom štádiu, ktoré sa normalizuje s vekom. Rôzne ranné urážky môžu túto normalizáciu u dospelých narušiť natoľko, že klasická os RAS je nevhodne aktivovaná, čo vedie k zvýšeniu TK a rozvoju ochorenia obličiek u dospelých potomkov. Okrem toho aberantná neonatálna supresia intrarenálneho RAAS prispieva k zmenám renálnej morfológie [9], čo je v súlade so štúdiami, ktoré uvádzajú blokádu RAAS ACEI alebo ARB [45].
Je pozoruhodné, že aberantná aktivácia RAAS môže byť transgeneračná. V modeli vysokofruktózovej diéty matky [99] sa zvýšenie TK pozorovalo u potomkov prvej a druhej generácie s maximálnym zvýšením hladín renínu, ANG II a aldosterónu v krvi u potomkov tretej generácie. Okrem toho príjem vysokej fruktózy matkou zvýšil renálnu expresiu mRNA ACE a AT1R počas viacerých generácií potomstva až do tretej. Bude narastať potreba lepšie pochopiť, či transgeneračná aktivácia RAAS má potenciálny vplyv na iné modely programovania obličiek.
3.4. Neklasická os RAAS v programovaní obličiek
Keďže neklasická os RAAS je vo všeobecnosti proti účinkom klasickej osi RAAS, predpokladá sa, že znížený tonus ACE2-ANG-(1-7)-MAS receptorového systému tiež prispieva k týmto patológiám . Podobne ako klasická os RAAS, aj neklasická os RAAS bola spojená s programovaním plodu [24]. Tabuľka 1 ukazuje, že u dospelých potomkov sa vyvinula hypertenzia a ochorenie obličiek, ktoré sa zhoduje so zníženou neklasickou dráhou RAAS v niekoľkých modeloch programovania obličiek, vrátane nízkobielkovinovej diéty matky [61], CKD u matky [71], diabetu matky [72] a modelu expozície glukokortikoidom [87,88]. Správy však boli v rozpore so zvýšenou expresiou ACE2 v modeli nepretržitej expozície svetlu [74].
3.5. Aldosterón v obličkovom programovaní
Aldosterón je hlavným regulátorom homeostázy sodíka. Izoforma 1 kinázy regulovanej sérom a glukokortikoidmi (SGK1) je kľúčovým mediátorom účinku aldosterónu v distálnom nefróne na reguláciu takmer všetkých sodíkových transportérov [100]. V porovnaní s inými zložkami v RAAS sa menej pozornosti venovalo hodnoteniu vplyvu aldosterónu na zvieracích modeloch programovania obličiek. Ako je uvedené v tabuľke 1, iba jedna správa preukázala, že hladina cirkulujúceho aldosterónu bola zvýšená u 8-týždňových potomkov narodených matkám vystaveným diéte s nízkym obsahom bielkovín [57]. Renálne sodíkové transportéry však boli študované na niekoľkých modeloch renálneho programovania, ako je prenatálne podávanie glukokortikoidov [84,101], nízkoproteínová diéta [58,102], model nepretržitého vystavenia svetlu [74] a kombinovaná diéta s vysokým obsahom fruktózy a soli [103]. ]. Rôzne ranné urážky ukázali, že programovanie obličiek je spojené so zvýšenými hladinami mRNA a množstvom proteínov niekoľkých sodíkových transportérov, ako je sodíkový výmenník vodíka typu 3 (NHE3), kotransportér Na-K-2Cl (NKCC2), Na plus / K plus podjednotka ATPázy a1 (NaKATPáza) a kotransportér Na plus/Cl- (NCC). Je pozoruhodné, že SGK1 môže byť aktivovaný glukokortikoidom a soľou, s výnimkou aldosterónu [104]. Preto, ak sú aberantné sodíkové transportéry vo vyššie uvedených zvieracích modeloch renálneho programovania priamo regulované aldosterónom alebo nečakajú na ďalšie objasnenie. Okrem toho objavujúce sa dôkazy ukazujú, že hypertenzia vyvolaná fruktózou súvisí s upreguláciou sodíkového transportéra NHE3 a predpokladaného aniónového transportéra 1 (PAT1) transportéra chloridov, aby sa stimulovala absorpcia sodíka a chloridu [105]. Keďže veľká časť predchádzajúcej práce skúmajúcej pôsobenie RAAS priamo študovala transportéry sodíka, bude potrebné lepšie pochopiť súhru medzi RAAS a transportérom chloridov pri hypertenzii.

púštne žijúce cistanche
4. Ústredná úloha RAAS pri sprostredkovaní spoločných mechanizmovProgramovanie obličiek
Vzhľadom na rôzne ranné urážky, ktoré vyvolávajú podobné renálne výsledky u dospelých potomkov, môžu existovať niektoré bežné mechanizmy patogenézy v obličkovom programovaní. Doteraz bolo identifikovaných niekoľko špecifických mechanizmov na vysvetlenie renálneho programovania. Tieto mechanizmy zahŕňajú aberantný RAAS, oxidačný stres, nedostatok oxidu dusnatého (NO), dysbiózu črevnej mikrobioty, dysregulované signály snímania živín, epigenetickú reguláciu a znížený počet nefrónov [6–10,13,18–20]. Je dôležité poznamenať, že medzi týmito navrhovanými mechanizmami je RAAS úzko spojený s ostatnými ako centrum pri určovaní procesov programovania obličiek. Vzájomné pôsobenie medzi RAAS a inými navrhovanými mechanizmami, ktoré sú základom renálneho programovania v reakcii na nepriaznivé inzulty v ranom veku, je znázornené na obrázku 3. Každý mechanizmus bude diskutovaný postupne.

4.1. Oxidačný stres
Ako je uvedené inde [106,107], kľúčovú úlohu oxidačného stresu, ktorý sa podieľa na hypertenzii a chorobách obličiek vývojového pôvodu, podporujú mnohé klinické a experimentálne štúdie. Nerovnováha obranného systému antioxidantov a produkcie reaktívnych foriem kyslíka (ROS) spôsobuje oxidačný stres, ktorý implikuje vývoj plodu [108]. Údaje z viacerých zvieracích modelov poukazujú na oxidačný stres zapojený do programovania obličiek [106,107]. Medzi nimi aberantný RAAS a oxidačný stres sú spojené s renálnym programovaním v modeloch prenatálnej expozície DEX [84], vysokofruktózovej diéty matky [53], diéty s vysokým obsahom tukov [66], CKD matky [71], preeklampsie [75 ], expozícia TCDD a dexametazónu u matky [82] a prenatálna DEX plus diéta s vysokým obsahom tukov po odstavení [86]. Je dobre známe, že ANG II pôsobiaci prostredníctvom AT1R je účinným aktivátorom NADPH oxidázy v obličkách natoľko, že zvyšuje produkciu ROS, čo sa podieľa na rozvoji hypertenzie [109]. Na druhej strane, zvýšenie AGT závislé od ROS hrá úlohu v progresii diabetickej nefropatie [110].
V modeli programovania obličiek sme pozorovali, že nevhodná aktivácia RAAS môže byť obnovená antioxidačnou terapiou [86]. Uvádza sa, že dimetylfumarát (DMF) aktivuje 2-faktor 2 odvodený od erytroidov (Nrf2, hlavný hráč v antioxidačnej obrane) a chráni pred poškodením oxidačným stresom [111]. Naša predchádzajúca práca ukázala, že podávanie DMF v tehotenstve chráni dospelých potomkov pred hypertenziou naprogramovanou prenatálnym DEX plus postnatálnou diétou s vysokým obsahom tukov, ktorá bola relevantná pre zníženú expresiu mRNA renínu, AGT, ACE a AT1R [86]. Hoci klinické štúdie využívajú induktory Nrf2 na liečbu CKD, aktivácia Nrf2 bola spojená s nepriaznivými účinkami, ako je proteinúria a nefrogénny diabetes insipidus [112, 113]. Do akej miery môže byť aktivácia Nrf2 prospešná pre CKD a ako sú Nrf2 a oxidačný stres prepojené s RAAS, sú otázky, ktoré čakajú na ďalšie objasnenie.
Ďalšia správa ukázala, že ochranné účinky melatonínu, silného antioxidantu, proti programovanej hypertenzii sa pripisujú zvýšeným renálnym hladinám ACE2 [75]. Okrem toho sme predtým skúmali model hypertenzie vyvolanej expozíciou svetla u matky a zistili sme, že terapia materským melatonínom chránila potomstvo pred hypertenziou, ktorá sa zhodovala so zvýšenou renálnou expresiou ACE2 [74]. Terapia melatonínom tiež zabránila zvýšeniu BP potomkov, ktoré sa zhodovalo so zvýšeným množstvom proteínu ACE2 v modeli kalorickej reštrikcie matky [114]. Tieto pozorovania naznačujú, že súhra medzi RAAS a oxidačným stresom sa podieľa na renálnom programovaní a následne nepriaznivých renálnych výsledkoch.
4.2. Nedostatok oxidu dusnatého
Značná pozornosť bola venovaná úlohe deficitu NO pri sprostredkovaní hypertenzie a ochorení obličiek vývojového pôvodu [19,115]. Jednou z hlavných príčin nedostatku NO je zvýšený asymetrický dimetylarginín (ADMA), endogénny inhibítor NOS [116]. Zameranie na dráhu ADMA/NO na zníženie ADMA a obnovenie NO sa považovalo za preprogramovací prístup na prevenciu programovania obličiek a následne hypertenzie a ochorenia obličiek [19,115].
ANG II môže znížiť biologickú dostupnosť NO podporou oxidačného stresu, zatiaľ čo NO je schopný vyvážiť vazokonstrikčný účinok ANG II [117]. V modeli expozície L-NG-Nitroarginín metylesteru matky (L-NAME, inhibítor NO syntázy) u matky deplécia NO spôsobila zvýšenie BP, ktoré sa zhodovalo so zvýšenou expresiou mRNA renínu a ACE v obličkách potomkov [75]. V inom modeli programovania obličiek blokáda RAAS aliskirenom chránila potomstvo dospelých potkanov pred hypertenziou naprogramovanou kalorickou reštrikciou matky v [118]. Ochranný účinok aliskirenu nie je nasmerovaný len na RAAS, ale aj prostredníctvom regulácie dráhy NO, reprezentovanej znížením plazmatických hladín ADMA a zvýšením hladín NOx v moči (NO2- plus NO3-) [ 118]. Podobne ako pri modeloch renálneho programovania sa uvádza, že skorá liečba aliskirenom blokuje rozvoj hypertenzie súvisiacej so znížením plazmatických hladín ADMA u spontánne hypertenzných potkanov (SHR), najčastejšie používaného modelu hypertenzie [119]. Keďže rovnováha medzi dráhou ADMA/NO a RAAS hrá rozhodujúcu úlohu v patogenéze programovania obličiek, bude narastať potreba lepšie porozumieť mechanizmom účinku RAAS na programovanie obličiek so zameraním na jeho súhru s NO. .
4.3. Znížený počet nefrónov
Deficit počtu nefrónov spôsobuje vysoký glomerulárny kapilárny tlak a glomerulárnu hyperfiltráciu, čo následne vedie k ďalšej strate nefrónov v neskoršom živote [8]. V súlade s tým sa nízky počet nefrónov považoval za životne dôležitý mechanizmus, ktorý je základom programovania obličiek. Niekoľko epidemiologických štúdií podporuje, že nízka pôrodná hmotnosť a predčasnosť, dva klinické náhradné markery počtu nefrónov, sú rizikovými faktormi pre hypertenziu a ochorenie obličiek v dospelosti [120–122]. U potkanov dospelí potomkovia vykazovali znížený počet nefrónov, keď sa DEX podával 2 dni v embryonálnych dňoch 13–14 alebo 17–18 [101]. Tieto zistenia naznačujú existenciu vývojových okien zraniteľnosti voči podmienkam prostredia počas vývoja obličiek. Ako sme už spomenuli, blokáda RAAS počas laktácie, neskorého štádia nefrogenézy u hlodavcov, vedie k zníženiu počtu nefrónov a hypertenzii v dospelosti [50].
Niekoľko zvieracích modelov programovania obličiek, ako je uvedené v tabuľke 1, naznačilo, že rôzne nepriaznivé vnútromaternicové stavy môžu viesť k nízkej výbave nefrónov a aberantnému RAAS súčasne, ako v prípade obmedzenia materského proteínu [56], expozície materským lipopolysacharidom (LPS) [76 ] a prenatálna hypoxia [78]. Prenatálna hypoxia viedla k zníženiu počtu nefrónov o 25 percent a zvýšeniu TK u potomkov dospelých samcov myší, čo súvisí so zvýšením renálnej expresie mRNA renínu (~2--násobne) a AT1R, ako aj koncentrácií renínu ( ~50 percentný nárast) [78].
Nízka zásoba nefrónov ako taká však nie je nevyhnutná pre hypertenziu a ochorenie obličiek vývojového pôvodu [8]. Úlohy RAAS pri zmene nefrónu
zdroj pre programovanie obličiek je stále potrebné identifikovať, ale je predmetom veľkého záujmu.
4.4. Epigenetická regulácia
Epigenetická regulácia je ďalším dôležitým mechanizmom, ktorý je základom fetálneho programovania [123]. Epigenetické mechanizmy pozostávajú z metylácie DNA, modifikácie histónov a nekódujúcich RNA (ncRNA). Globálne vzorce metylácie DNA v niekoľkých orgánoch boli hodnotené v rôznych modeloch vývojového programovania, ako je nízkoproteínová diéta matky [124], fajčenie matky [125] a nedostatok mikroživín [126]. Menej pozornosti sa však venovalo obličkám. Aberantná metylácia DNA bola spojená s hypertenziou vývojového pôvodu [127]. Pri SHR je zvýšená expresia AT1R relevantná pre progresívnu hypometyláciu v promótore AT1R, keď sa hypertenzia objaví vo veku 20 týždňov [128]. Uvádza sa však, že gén AT1R je hyper- alebo hypometylovaný v rôznych modeloch programovanej hypertenzie [23,129].
Okrem toho dochádza k epigenetickej modifikácii histónu, keď je N-terminálny koniec vystavený rôznym posttranslačným modifikáciám [130]. Jednou z najčastejších epigenetických modifikácií je acetylácia histónov, ktorá je katalyzovaná histónacetyltransferázami (HAT). Naopak, históndeacetylázy (HDAC) určujú deacetyláciu histónu. Presluchy medzi HDAC a RAAS boli navrhnuté na riadenie rozvetvenia uretických púčikov počas vývoja obličiek [18]. HDAC boli hlásené pre regulovanú expresiu niekoľkých génov patriacich do RAAS, vrátane AGT, renínu, ACE a AT1R [131]. Naša predchádzajúca štúdia ukázala, že trichostatín A, inhibítor HDAC, zabránil neonatálnej DEX-indukovanej programovanej hypertenzii sprevádzanej znížením AGT, ACE a ACE2 [87].
ncRNA sa podieľajú na niekoľkých epigenetických procesoch [132] a mikroRNA (miRNA) sú najčastejšie študované malé ncRNA. Pokiaľ ide o gény regulované RAAS, analýza miRNA väzbových miest pomocou TargetScan [133] naznačila, že 368 rôznych rodín miRNA sa zameriava na prvky RAAS, z ktorých väčšina zdieľa transkripty. V modeli reštrikcie materského proteínu bol prechod obličkového epitelu na mezenchým spojený so zníženou hladinou miR-200a, miR-141 a miR-429 [134]. Ďalšia správa preukázala, že mmu-miR-27a a mmu-miR{13}}b upregulovali ACE, zatiaľ čo mmumir{14}} downregulovali AT2R u potomkov narodených matkám s obmedzením bielkovín [135]. Jedna miRNA však môže regulovať množstvo mRNA, čo sťažuje dešifrovanie presných mechanizmov zapojených do programovania obličiek. Na objasnenie presnej povahy mechanizmov a na vývoj potenciálnych terapeutických aplikácií sú potrebné ďalšie štúdie na ľuďoch a experimentálne štúdie.
4.5. Iní
Existujú aj ďalšie hlásené mechanizmy za programovaním obličiek, pomocou ktorých môže RAAS pôsobiť: (1) dysregulované signály snímania živín, (2) dysbióza črevnej mikroflóry a (3) rozdiely medzi pohlaviami. Po prvé, nutričné urážky v ranom veku môžu zhoršiť signály vnímania živín, ktoré ovplyvňujú vývoj plodu a následne programujú hypertenziu v neskoršom veku [136]. Receptor aktivovaný peroxizómovým proliferátorom (PPAR), jeden zo signálov snímajúcich živiny, môže byť sprostredkovaný inými signálmi snímajúcimi živiny na reguláciu expresie cieľových génov PPAR [137]. Je potrebné poznamenať, že niekoľko cieľových génov PPAR patrí ku komponentom RAAS alebo sodíkovým transportérom, ako je renín a SGK1 [138]. Ako je uvedené inde [138], objavujúce sa dôkazy naznačujú, že včasná intervencia modulátormi PPAR môže zabrániť hypertenzii vývojového pôvodu. Preto sa špekuluje, že RAAS môže interagovať so signálmi snímania živín na programovanie hypertenzie a ochorenia obličiek.
Po druhé, nepriaznivé vnútromaternicové stavy môžu narušiť črevnú mikrobiálnu rovnováhu, čo má za následok následné nepriaznivé výsledky u potomstva vrátane hypertenzie [139]. Predchádzajúci výskum ukázal, že ACE2 má nekatalytickú úlohu v biológii čreva a moduluje zloženie črevnej mikroflóry [140]. Keďže dysbióza črevného mikrobiómu bola spojená s hypertenziou moduláciou črevného RAAS [141], tieto zistenia naznačujú, že medzi črevnou mikroflórou a RAAS môže existovať vzťah, ktorý je základom patogenézy programovania obličiek, hoci to zostáva špekulatívne.
Napokon, objavujúce sa dôkazy podporujú rozdiely v závislosti od pohlavia, pokiaľ ide o hypertenziu a ochorenie obličiek vývojového pôvodu [142,143]. Je pozoruhodné, že RAAS bol hlásený ako reakcia špecifická pre pohlavie na environmentálne urážky [144]. Tiež zmena v odpovedi renálneho transkriptómu na rôzne inzulty je závislá od pohlavia [52,145,146]. Väčšina zvieracích modelov obličkového programovania, ako je uvedené v tabuľke 1, však vyšetruje iba mužov namiesto oboch pohlaví. Bude teda narastať potreba objasniť vplyv RAAS na mechanizmy závislé od pohlavia za programovaním obličiek a byť schopný vyvinúť nové stratégie špecifické pre pohlavie zamerané na RAAS na prevenciu programovanej hypertenzie a ochorenia obličiek vývojového pôvodu u oboch pohlaví.
Hoci viaceré mechanické väzby uvedené vyššie, RAAS funguje ako centrálne spojenie pre hypertenziu a ochorenie obličiek vývojového pôvodu. Lepšie pochopenie interakcie medzi RAAS a inými spoločnými mechanizmami, ako aj zameranie sa na RAAS s cieľom vyvinúť intervenciu preprogramovania, sú kľúčom k včasnej prevencii alebo liečbe prehypertenzie a subklinického ochorenia obličiek.

plátky cistanche tubolosa
5. Zameranie na RAAS ako stratégie preprogramovania
Stratégie preprogramovania zamerané na RAAS na prevenciu vývojového programovania hypertenzie a ochorenia obličiek, ktoré boli použité na rôznych zvieracích modeloch, sú uvedené v tabuľke 2 [52, 63, 118, 119, 147 – 154]. V súčasnosti bolo popísaných niekoľko terapeutických zásahov, ako je inhibítor renínu [52,118,119], ACEI [63,147–150], ARB [118,151,152], AT1R antisense [153] a aktivátor ACE2 [154]. Hlavné ochranné účinky rôznych intervencií založených na RAAS na nepriaznivé renálne výsledky sú proti hypertenzii [52,63,118,119,147–154], po ktorej nasleduje albuminúria [149], renálna dysfunkcia [150] a renálna fibróza [154]. Účinky preprogramovania terapií založených na RAAS sa skúmali u potkanov vo veku od 9 týždňov do 6 mesiacov, čo je takmer ekvivalent ľudského veku od detstva po mladú dospelosť. Väčšina štúdií sa však zamerala len na mužov a netestovali rôzne dávky. Či sa tieto pozorované účinky objavia v závislosti od dávky alebo pohlavia, čaká na objasnenie ďalších štúdií.
Včasná blokáda klasickej osi RAAS bola navrhnutá na preprogramovanie nevhodne aktivovaného RAAS na prevenciu hypertenzie a ochorenia obličiek vývojového pôvodu.
Liečba malých potomkov inhibítorom renínu aliskirenom [52,118], ACEI kaptoprilom [63] alebo ARB losartanom [118,151] vo veku 2–4 týždňov sú najbežnejšími terapeutickými obdobiami na kompenzáciu účinkov programovania vývoja na BP.
K dnešnému dňu je aliskiren jediným inhibítorom renínu schváleným na liečbu hypertenzie. Aliskiren však nemôže zabrániť interakcii medzi PRR a jeho ligandom. Aj keď boli na zvieracích modeloch hlásené priaznivé účinky pre PRR inhibičný peptid, peptid manipulačnej oblasti a PRO20 [155, 156], účinnosť týchto peptidov v špecifickosti je otázna [94]. Dúfame teda, že navrhnutie špecifického nepeptidového inhibítora PRR by mohlo v blízkej budúcnosti viesť k priaznivej inhibícii (pro)renínu-PRR.
Všetky predchádzajúce práce skúmajúce zásahy do preprogramovania skúmali iba potkany. Keďže nefrogenéza je u potkanov dokončená v druhom postnatálnom týždni, takmer všetky intervencie blokády RAAS na prevenciu hypertenzie a ochorenia obličiek začínajú už dva týždne po narodení. Hoci sa antisense podanie AT1R u SHR uskutočnilo na 5. deň po narodení [153], jeho vplyv na počet nefrónov sa zatiaľ neskúmal.
Odhliadnuc od klasickej osi v RAAS, nové dôkazy poskytujú ochrannú úlohu neklasickej osi pri diagnostikovanej hypertenzii a ochorení obličiek, čo pripravuje pôdu pre nové terapeutické prístupy [21]. Napriek tomu sa málo pozornosti venovalo aplikácii tohto prístupu na programovanú hypertenziu a ochorenie obličiek. Podľa tabuľky 2 iba jedna štúdia uvádza, že podávanie s diminazén aceturátom (DIZE), predpokladaným aktivátorom ACE2, alebo s ANG -(1–7) počas gravidity môže zmierniť hypertenziu a renálnu fibrózu u dospelých SHR potomkov [154]. Vzhľadom na aktiváciu osi ACE2-ANG-(1-7)-MA s terapeutickým potenciálom pri diagnostikovanej hypertenzii a ochorení obličiek pretrváva potreba ďalšej štúdie na objasnenie jej preprogramovacích účinkov na programovanie obličiek. V literatúre chýba hlbšie pochopenie toho, ktorá zložka RAAS je pre cielený prístup najdôležitejšia a aký čas je optimálnym terapeutickým oknom na prevenciu hypertenzie a ochorení obličiek vývojového pôvodu.

6. Závery
Súčasné dôkazy poskytli rázne, ale neúplné údaje, pokiaľ ide o potenciálnu terapeutickú úlohu intervencií založených na RAAS pri hypertenzii a chorobách obličiek vývojového pôvodu. Tento prehľad poskytuje stručný prehľad rôznych terapií založených na RAAS, ktoré ukazujú výhody programovania obličiek, vrátane inhibítora renínu, ACEI, ARB, AT1R antisense a aktivátora ACE2.
Doteraz jedným z hlavných nevyriešených problémov je, že takmer žiadne štúdie neprijali holistický prístup na súčasné kvantifikovanie expresie/aktivity celého repertoáru komponentov RAAS v experimente. Vzhľadom na komplexnú povahu signalizácie RAAS je účinok preprogramovania v reakcii na intervencie založené na RAAS v ranom veku, či už jednotlivo alebo v kombinácii, neúplný a ťažko predvídateľný. Preto je potrebná budúca práca na vývoji ideálnej metodológie, aby sme získali holistickejší pohľad na RAAS a zabezpečili, že terapia založená na RAAS sa bude uplatňovať len správnym smerom. Okrem toho bude potrebné venovať pozornosť rozhodnutiu o optimálnej dávke v závislosti od pohlavia, aby sa maximalizoval prínos bez zvýšenia toxicity pred klinickým prekladom.
Napriek výraznému pokroku dosiahnutému v dostupnosti širokej škály liekov na báze RAAS sa menej pozornosti venovalo skúmaniu ich preprogramovacích účinkov na hypertenziu a ochorenie obličiek. Ďalšou výzvou je, že špecifické vývojové okná pre rôzne terapie založené na RAAS na preprogramovanie procesov poháňajúcich hypertenziu a ochorenie obličiek stále čakajú na ďalšie objasnenie.
Náš prehľad zatiaľ urobil krok vpred tým, že prepojil RAAS s hypertenziou a ochorením obličiek vývojového pôvodu, čo môže priniesť pohľad na nové intervencie založené na RAAS na prevenciu porúch súvisiacich s programovaním obličiek v klinickom prostredí.

extrakt z cistanche tubolosa: zlepšuje funkciu obličiek
Príspevky autora:C.-NH: podieľal sa na tvorbe koncepcie, koncipovaní rukopisu, kritickej revízii rukopisu, interpretácii údajov a schválení článku; Y.-LT: podieľal sa na návrhu rukopisu, interpretácii údajov, generovaní koncepcie, kritickej revízii rukopisu a schválení článku. Všetci autori si prečítali publikovanú verziu rukopisu a súhlasili s ňou.
Financovanie:Tento výskum bol financovaný nemocnicou Chang Gung Memorial Hospital, Kaohsiung, Taiwan, grantmi CMRPG8J0251, CMRPG8J0252, CMRPG8J0253, CMRPG8J0891 a CMRPG8J0892.
Vyhlásenie inštitucionálnej revíznej rady:Nepoužiteľné.
Vyhlásenie informovaného súhlasu:Nepoužiteľné.
Konflikt záujmov:Autori nedeklarujú žiadny konflikt záujmov. Investori nezohrávali žiadnu úlohu pri navrhovaní štúdie; pri zbere, analýzach alebo interpretácii údajov; v písaní rukopisu, alebo v rozhodnutí o zverejnení výsledkov.
1 Department of Pharmacy, Kaohsiung Chang Gung Memorial Hospital, Kaohsiung 833, Taiwan;cnhsu@cgmh.org.tw
2 Farmaceutická fakulta, Kaohsiung Medical University, Kaohsiung 807, Taiwan
3 Oddelenie pediatrie, nemocnica Kaohsiung Chang Gung Memorial Hospital a Univerzita Chang Gung University College of Medicine, Kaohsiung 833, Taiwan
4 Inštitút pre translačný výskum v biomedicíne, Kaohsiung Chang Gung Memorial Hospital a ChangGung University College of Medicine, Kaohsiung 833, Taiwan
Referencie
1. Svetová zdravotnícka organizácia. Hypertenzia. 2019. Dostupné na internete: https://www.who.int/news-room/fact-sheets/detail/ hypertension(prístup 12. januára 2021).
2. Lozano, R.; Naghavi, M.; Foreman, K.; Lim, S.; Shibuya, K.; Aboyans, V.; Abrahám, J.; Adair, T.; Aggarwal, R.; Ahn, SY; a kol. Globálna a regionálna úmrtnosť z 235 príčin smrti pre 20 vekových skupín v rokoch 1990 a 2010: Systematická analýza pre štúdiu Global Burden of Disease 2010. Lancet 2012, 380, 2095–2128.
3. Weir, MR Hypertenzia a oblička: Pohľady na vzťah obličkových chorôb a kardiovaskulárnych chorôb. Clin. J. Am. Soc. Nephrol. 2009, 4, 2045–2050.
4. Herrmann, SM; Textor, SC Renovaskulárna hypertenzia. Endocrinol. Metab. Clin. N. Am. 2019, 48, 765–778.
5. Basso, N.; Terragno, NA História o objave systému renín-angiotenzín. Hypertenzia 2001, 38, 1246–1249.
6. Zandi-Nejad, K.; Luyckx, VA; Brenner, BM Hypertenzia dospelých a ochorenie obličiek: Úloha programovania plodu. Hypertenzia 2006, 47, 502-508.
7. Chong, E.; Yosypiv, IV Vývojové programovanie hypertenzie a ochorenia obličiek. Int. J. Nephrol. 2012, 2012, 760580.
8. Tain, YL; Hsu, CN Vývojový pôvod chronického ochorenia obličiek: Mali by sme sa zamerať na skorý život? Int. J. Mol. Sci. 2017, 18, 381.
9. Kett, MM; Denton, KM Renálne programovanie: Dôvod na obavy? Am. J. Physiol. Regul. Celé číslo. Comp. Physiol. 2011, 300, R791 – R803.
10. Paixao, AD; Alexander, BT Ako ovplyvňuje obličky perinatálne prostredie matky na rozvoj hypertenzie. Biol. Reprod. 2013, 89, 144.
11. Barker, DJ Fetálne programovanie koronárnej choroby srdca. Trendy. Endocrinol. Metab. 2002, 13, 364-368.
12. Haugen, AC; Schug, TT; Collman, G.; Heindel, JJ Evolúcia DOHaD: Vplyv environmentálnych vied o zdraví. J. Dev. Orig. Health Dis. 2015, 6, 55–64.
13. Tain, YL; Joles, JA Preprogramovanie: Preventívna stratégia pri hypertenzii so zameraním na obličky. Int. J. Mol. Sci. 2016, 17, 23.
14. Paauw, ND; van Rijn, BB; Lely, AT; Joles, JA Tehotenstvo ako kritické okno pre reguláciu krvného tlaku u matky a dieťaťa: Programovanie a preprogramovanie. Acta Physiol. 2017, 219, 241–259.
15. Lee, RM; Borkowski, KR; Leenen, FH; Tsoporis, J.; Coughlin, M. Kombinovaný účinok neonatálnej sympatektómie a demodulácie nadobličiek na krvný tlak a vaskulárne zmeny u spontánne hypertenzných potkanov. Circ. Res. 1991, 69, 714-721.
16. Vavrinovd, A.; Behuliak, M.; Bencze, M.; Vodička, M.; Ergang, P.; Vaněčkovd, I.; Zicha, J. Sympatektómiou indukované zníženie krvného tlaku u dospelých normotenzných a hypertenzných potkanov je ovplyvnené zvýšenou kardiovaskulárnou citlivosťou na vazokonstriktory. Hypertenzia. Res. 2019, 42, 1872–1882.
17. Paz Ocaranza, M.; Riquelme, JA; Garcia, L.; Jalil, JE; Chiong, M.; Santos, RAS; Lavandero, S. Protiregulačný systém renín-angiotenzín pri kardiovaskulárnych ochoreniach. Nat. Rev. Cardiol. 2020, 17, 116–129.
18. Bagby, SP Výživa matky, nízky počet nefrónov a hypertenzia v neskoršom veku: Cesty nutričného programovania. J. Nutr. 2007, 137, 1066–1072.
19. Hsu, CN; Tain, YL Regulácia produkcie oxidu dusnatého vo vývojovom programovaní hypertenzie a ochorenia obličiek. Int. J. Mol. Sci. 2019, 681.
20. Hsu, CN; Tain, YL Programovanie v ranom veku a preprogramovanie ochorenia obličiek a hypertenzie u dospelých: Súhra medzi výživou matiek a oxidačným stresom. Int. J. Mol. Sci. 2020, 21, 3572.





